mr peabody
Bluelight Crew

Alexander Shulgin
An Introduction to Psychedelic Neuroscience
Tanya Calvey, Fleur Margaret Howells | January 2018
This chapter is an introduction to the volume Psychedelic Neuroscience of Elsevier's Progress in Brain Research addressing the neurobiological mechanisms of psychedelic drugs, the resulting changes in brain activity and integration of traditional viewpoints. As the field is relatively new, there are discrepancies in the literature related to classification, composition and effects of the various psychedelics. Currently, psychedelics are grouped according to their neuro-receptor affinities into classic and atypical psychedelics, each with individual treatment potentials and abilities to elicit potent acute experiences and long-lasting changes in neurobiology through concurrent activation of several neuromodulatory systems. There is disparity in psychedelic brain imaging studies, delineating what is neural activity and hemodynamic needs further investigation for us to understand the brain state changes that are apparent. The psychedelic brain state is often compared to acute psychosis and we review the psychedelic animal models of psychosis and human brain imaging studies and contrast these to psychosis. The term psychedelic means mind-revealing and psychedelics have exceptional anti-amnesic effects and are able to make conscious that which was previously unconscious through changes in brain state, but also there is growing evidence which demonstrates the role of epigenetic mechanisms. This supports traditional therapeutic use of psychedelics to heal ancestral trauma. Details of these mechanisms are provided along with suggestions for further research.
WHAT ARE PSYCHEDELICS?
The field of psychedelic neuroscience has witnessed a recent renaissance following decades of restricted research due to their legal status. As this is a relatively new field, there are incongruences in the literature related to terminology, classification, content and effect of the various psychedelics.
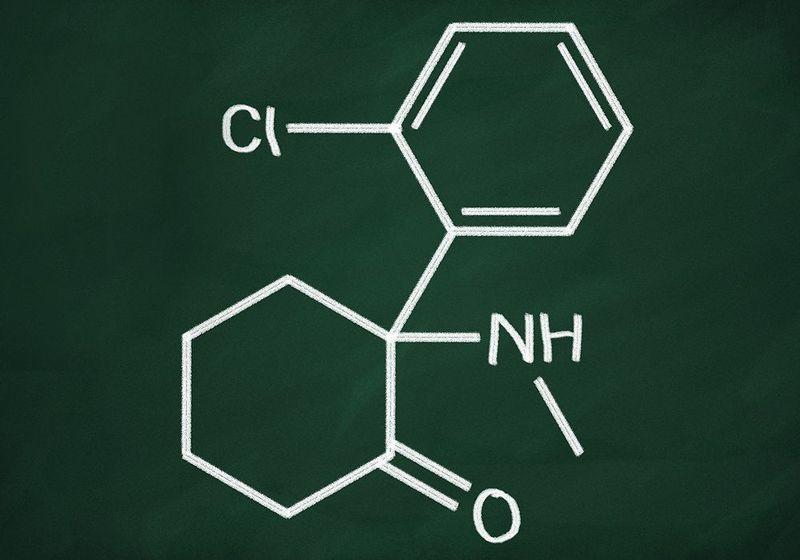
The currently accepted classification of psychedelics includes classic psychedelics and atypical/non-traditional/non-classical psychedelics. Classic psychedelics are the phenethylamines such as mescaline, tryptamines such as 5-MeO-DMT, DMT, psilocybin, and ergolines such as LSD. Atypical psychedelics can be further divided into dissociative psychedelics, e.g., PCP, ketamine and ibogaine, as well as cannabinoid agonists (e.g. THC), muscarinic receptor antagonists (e.g., scopolamine), and entactogens (e.g., MDMA).
Problematic in the field, perhaps more so than in other fields, is the issue of conflicting results, likely due to the limited research. For example, there is conflicting evidence as to whether or not the low 5HT2A affinity of ibogaine has any functional relevance. Ly et al. found that the 5HT2A receptor antagonist, ketanserin, blocked the effect of noribogaine on structural plasticity yet both ibogaine and noribogaine failed to induce head-shake response in rats, a behavior which is seen to be comparable to hallucinations in humans and mediated by 5HT2A activation. Gonzalez et al. suggest that this result supports the subjective experiences in humans, where ibogaine does not produce the typical interferences in thinking, identity distortions, and space-time alteration, which are produced by the classic psychedelics. A second example, where there is confusion is the literature regarding 5-MeO-DMT. 5-MeO-DMT is described as being a constituent of ayahuasca, while it is evidenced that ayahuasca holds a high concentration of DMT, 5-MeO-DMT concentration is either non-existent or negligible in most brews, and the ayahuasca psychedelic experience bears little to no resemblance to an experience with 5-MeO-DMT. Then cannabis research is possibly the best example of conflicting psychedelic research. As reviewed in Colizzi and Bhattacharyya, evidence as to whether cannabis use is associated with adverse effects to mental health or cognition in humans is equivocal. There are also many divergent results in the functional human neuroimaging studies as detailed by Meuller et al. Indeed, this is a new and growing field of research and more research is required to uncover the various psychedelics drugs, their active components and neurobiological effects.
NEUROBIOLOGY OF PSYCHEDELIC THERAPY FOR DEPRESSION AND ADDICTION
Classic psychedelics and dissociative psychedelics are known to have rapid onset antidepressant and anti-addictive effects, unlike any currently available treatment. Randomized clinical control studies have confirmed antidepressant and anxiolytic effects of classic psychedelics in humans. Ketamine also has well established antidepressant and anti-addictive effects in humans mainly through its action as an NMDA antagonist. Ibogaine has demonstrated potent anti-addictive potential in pre-clinical studies and is in the early stages of clinical trials to determine efficacy in robust human studies.
Psychedelics are not only known to have rapid onset but their effects persist long after their acute effects; this includes changes in mood and brain function. These effects are suggested to result from their unique receptor affinities which affect neurotransmission via neuromodulatory systems which then serve to modulate brain activity, i.e., neuro-plasticity. These lasting effects are reported to promote cell survival, be neuroprotective, and modulate neuroimmune systems of the brain. The mechanisms which lead to these long-term neuromodulatory changes have been linked to epigenetic modifications and gene expression changes. These psychedelic drug effects, previously under-researched, may potentially provide the next-generation of neurotherapeutics, where treatment resistant diseases, e.g., depression and addiction, may become treatable with attenuated pharmacological risk profiles.
Classic psychedelics have been shown to stimulate the serotonergic system mainly via 5HT1A, 5HT2A, 5HT2C, and 5HT7 receptors, with the dopaminergic system primarily via D2 receptors, and indirectly with the glutamatergic and GABAergic systems. There is significant cross-talk between these neuromodulatory systems and classic psychedelics, and activation of 5HT1A and 5HT2A receptors modulates glutamatergic and dopaminergic neurotransmission in brain networks associated with depression and addiction. Then activation of 5HT2C receptors localized in dopaminergic and GABAergic neurons in the ventral tegmental area (VTA) regulates motivation by modulating transmissions to the nucleus accumbens (NAc) and altered balance in this 5HT2C receptor-associated network is postulated to cause reward-related disorders, such as schizophrenia, depression, and addiction. Further, it is acknowledged that classic psychedelics have an extremely low potential for abuse, and it is suggested that stimulation of 5HT2C receptors limits their potential for addiction and that their therapeutic effects are mediated by acute 5HT2C receptor stimulation followed by sustained downregulation of 5HT2A and 5HT1A receptors. Investigation into the complex mechanisms of action of classic psychedelics which lead to its anti-depressant and anti-addiction properties via the serotonergic system continues to gain momentum, e.g., minimal research has investigated the role of 5HT7 activation by psychedelics, but has been suggested to play a role in classic psychedelic anti-addiction properties, specifically 5-MeO-DMT in alcohol use disorder.
The classic psychedelics act directly via the serotonergic system whereas certain atypical psychedelics have direct affinity for numerous neuromodulatory systems. A remarkable example, ibogaine, addressed by Corkery and Barsuglia et al., demonstrates novel pharmacological mechanisms of action to be considered in the potential treatment of substance use disorders and depression through simultaneous activation of multiple neuro-transmitter systems. This alkaloid has low micromolar affinity for mu and kappa opioid receptors, SIGMA-1 and SIGMA-2 receptors, serotonin reuptake transporter (SERT) and dopamine transporter (DAT), is an antagonist to NMDA and α3β4 nicotinic acetylcholine (nAChR) receptors and a weak 5HT2A receptor agonist. Drugs with similar NMDA affinity (e.g., ketamine and memantine) or NMDA regulators have shown promise in reducing symptoms of substance use disorders and depression. The mu-opioid receptor has demonstrated a functional role in drug reward as well as craving. Further, ibogaine possesses an opiate replacement mechanism of action as reported for compounds such as methadone. However, neither ibogaine nor its principle psychoactive metabolite, noribogaine, activate G-proteins associated with morphine administration, or produce signs and symptoms of opioid intoxication in opioid naive persons; therefore, it seems that ibogaine is able to produce a neuroadaptive effect on endogenous opioid systems which reverses opi- oid tolerance and may be implicated broadly in its addiction-interrupting effects.
ADDITIONAL THERAPEUTIC MECHANISMS OF ACTION
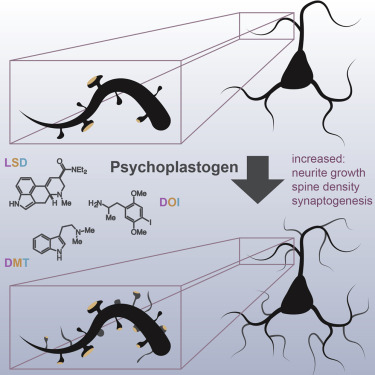
There is strong evidence suggesting that various psychedelics influence the expres- sion and modulation of genes, which in turn, may lead to long-term neurochemical and neuroplastic changes and modification of epigenetic mechanisms. When administered chronically, LSD has been shown to have effects on dopaminergic neurotransmission at the level of gene expression by decreasing the mRNA expression of the dopamine receptor genes DRD1 and DRD2 in the medial prefrontal cortex (mPFC). Classic and dissociative psychedelics lead to structural and functional changes in cortical neurons, with plasticity-promoting properties that rival brain-derived neurotropic factor (BDNF). LSD has demonstrated to be extremely potent in this regard possibly due to slow off kinetics of the LSD-bound 5HT2B crystal structure. Although the molecular targets of classic and dissociative psychedelics differ, their plasticity promoting properties are similar and are known to activate TrkB, mTOR, and 5HT2A signaling pathways, suggesting that these key signaling hubs may serve as potential targets for the development of psychoplastogens, fast-acting antidepressants, and anxiolytics. The mTOR signaling pathways may also play a role in certain psychedelics' ability to modify epigenetic mechanisms as mTORC1 is thought to be involved in regulating gene expression through epigenetic mechanisms or by directly affecting RNA stability/degradation. Another relevant receptor involved in epigenetic modification is SIGMAR1. DMT and ibogaine both activate SIGMAR1 and recently, SIGMAR1 has been shown to modulate epigenetic processes by creating a dose-dependent interaction between emerin and histone deacetylase (HDAC)1, HDAC2 and HDAC3, affecting chromatin compaction and gene expression. At the nuclear envelope, SIGMAR1 recruits chromatin-remodeling molecules to regulate gene expression and at the synaptic level it interacts with voltage-gated ion channels, which leads to modification and reorganization of several homo- and heteroreceptor complexes which in turn modulates of neurotransmission, reviewed in Inserra.

NEUROBIOLOGY OF THE PSYCHEDELIC EXPERIENCE
The psychedelic experience can produce eyes-closed and eyes-open imagery. The imagery can be clear and visual or it can have a dream-like quality with strong emotions and insights. There is also often heightened memory retrieval where long-forgotten memories are retrieved with exceptional clarity and detail. A common experience on ibogaine, for example, is that one's life flashes before their eyes as clear as if they were watching a movie about their life.
The neurophysiological mechanisms that facilitate the psychedelic experience are largely unknown but progress is being made in understanding how the neurochemical profile of psychedelics elicits this effect, bringing that which was previously unconscious to the conscious mind. The differences in the psychedelic state elicited by the various classic and atypical psychedelics hold promise in differentiating the exact neuroreceptor-mind interactions. Neuroreceptors are coded by individual genes and extensive genetic variation exists in the population for these neuromodulatory systems. Variation in the neuroreceptor-mind interactions, thus, holds further promise for psychiatric neurogenomics research as it will uncover how individual genetic variation in the neuromodulatory systems affects different psychedelic states and changes in neurobiology.
The serotonergic system's involvement in the psychedelic state has received the most research attention, and has provided new insights related to the neurochemical mechanisms underlying changes in brain network activity and perfusion. Serotonin is involved in many neurological (e.g., epilepsy) and psychiatric (e.g., depression) diseases. Serotonin receptors may directly or indirectly depolarize or hyperpolarize neurons by changing the ionic conductance and/or concentration within the cells and is able to change excitability within brain networks. For example, pharmacological magnetic resonance imaging (phMRI) in rats indicates that psilocin induces brain signal increases in olfactory and limbic areas and brain signal decreases in somatosensory and motor cortices. 5-MeO-DMT disrupts cortical activity and low frequency cortical oscillations in the frontal cortex of rats with alternating activity in frontal and visual areas associated with psychedelic effects.
In humans, activation of postsynaptic 5HT2A receptors in layer V of the medial prefrontal cortex mPFC is considered to be responsible for the visual hallucinations produced by classic psychedelics. Cortical 5HT2A hyper-activation affects cortico-striatal-thalamo-cortical circuit functioning and triggers a disruption in the thalamic gating of sensory and cognitive information leading to perceptual distortions.
Studies point toward not only brain network activity changes but also significant increases in hemodynamics. Increased cerebral blood flow has been reflected in temperature record of different brain areas with administration of MDMA, increased blood flow to the cortex correlated with increased neuronal activity as expected, however within the thalamus increased blood flow negatively correlated with increased neuronal activity. Then a psilocin study reported decreased local field potentials to sensory stimuli while hemodynamic response was enhanced. These data support a brain state change, but also require us to challenge our knowledge of the nature of this differential activity, neural versus hemodynamic, and their interplay which leads to the psychedelic experience.
Human brain imaging studies are limited, the neurophysiological underpinnings are largely speculative, while the findings are interesting and need to be further investigated. For example, an LSD functional MRI (fMRI) study found increased hemodynamic activity within brain areas rich in 5HT2A receptors and globally, the authors conclude that this reflected increased functional connectivity and that this increased activity led to ego dissolution. Then a psilocybin fMRI study found decreased hemodynamic activity within the thalamus and anterior and posterior cingulate cortices, where the decreased activity in the anterior cingulate correlated with the psychedelic experience. They further report a decrease in resting state connectivity between the medial prefrontal cortex and posterior cingulate cortex; the authors conclude that psilocybin reduced connectivity, and this reduction enabled the psychedelic experience. A different psilocybin resting state fMRI study reported that connectivity increased in higher brain networks such as the default mode, executive control, and dorsal attention networks. Then an arterial spin labelling study found that post MDMA, hemodynamics decreased and this was localized to right medial temporal lobe (MTL), thalamus, inferior visual cortex, and somatosensory cortex. Then a spontaneous magnetoencephalo-graphic (MEG) study investigated Lempel-Ziv (LZ) complexity, more commonly applied in EEG studies, to determine diversity of mixed-signal being recorded. Where they found three psychedelic drugs: psilocybin, LSD, and ketamine, increased signal diversity within occipital cortices, this extended over the parietal cortex with LSD, and even further with ketamine over the full cortex, excluding medial frontal brain areas, they were also able to support a reduction in alpha MEG activity for all three psychedelics tested.
The authors concluded that their findings provide confirmation that psychedelics create a higher level of consciousness which is reflected in MEG LZ complexity. A second psychedelic MEG study, which investigated psilocybin, reported reduction in spontaneous cortical activity in posterior association cortices, and in frontal association cortices. Second, they reported hugely significant reductions within the default mode network. Then a low-resolution electromagnetic tomography (LORETA) study investigated ayahuasca and found reductions in delta, theta, alpha-2, and beta-1 frequency bands, these changes were predominantly evident over the temporo-parietal-occipital junction. Overall, the brain imaging studies, which address hemodynamic activity in humans, support acute increases and decreases in several brain regions. Electro-magnetic studies report reductions in neural activity, most notably evident for alpha band frequency, and not limited to specific brain areas, but potentially reducing frontal activity and increasing parietal activity. Together these findings support the conclusion Lebedev et al. drew in relation to psilocybin; that psychedelics lead to a disintegration of functional connectivity, and this permits ego-dissolution. It is suggested that this disintegration is at least in part due to decreased connectivity between of the parahippocampal and retrosplenial cortex, as was reported in a resting state fMRI LSD study, which may serve to support the change in cortical activity.
Psychedelic electroencephalographic research supports a brain state change, with evidence of reduced neural activity frontally and support increased activity parietally. A quantitative EEG (qEEG) study, investigated the acute effects of DMT and 5-MeO-DMT, finding significant reductions in global absolute alpha activity, while moderate significant increases were seen in theta and beta. Then ayahuasca induced decreases in delta, theta, and alpha frequency activity, the power of alpha activity in parietal and occipital cortex was negatively related with the intensity of visual psychedelic experience. Acute dosing of DMT showed reduction in coherence between anterior and posterior EEG recording sites, where anterior coherence decreased, permitting parietal coherence to drive the EEG activity. Psilocybin was reported to decrease 1.5-20 Hz frequency activity, and using source density EEG the neural networks which showed this decreased connectivity were in the anterior and posterior cingulate cortices and the parahippocampal regions. The authors also report that the psychedelic experience was related to delta activity between the retrosplenial cortex, the parahippocampus, and the lateral orbitofrontal area. These data together support shifts in brain state with changes in cortical control from frontal brain areas to parietal and in part this is related to hippocampal network activity.
Further, EEG studies have investigated and contrasted MDMA, cannabis, and MDMA with cannabis use on EEG frequency where they show that MDMA with or without cannabis use increased delta band frequency, while cannabis use alone was found to increase high alpha band activity. In addition, this study found that MDMA with or without cannabis use showed increased blood flow and diastolic blood velocity. The increased alpha band activity in cannabis-alone users contradicts several papers which support attenuated alpha band activity. Then other cannabis studies report no change in alpha, and only report reductions in delta and beta activity. Cannabis has been shown to increase heart rate when alpha band activity is reduced, CB1 antagonism prevents the increase in heart rate produced by cannabis; however, this study which tested CB1 antagonism in acute cannabis use reported no change in alpha activity. Then an acute study investigated the effects of MDMA with and without ethanol or THC, and MDMA alone was found to decrease theta and alpha power, when a combination of MDMA and THC was taken together their attenuation of theta and lower-1-alpha was less than when given alone, then the combination of MDMA with THC lead to significant reduction in lower-2-alpha, but not when administered alone. Then a study in users of MDMA showed an incremental increase in alpha activity as cumulative doses of ecstasy increased, with no effect on theta activity.
There is an absence of reliable ibogaine EEG studies in humans, a single study in cynomologus monkeys reported no effect, then several rat studies. These studies report increased low-frequency, delta and theta, activity, and found ibogaine pre-treatment lowered cocaine-induced seizure threshold, reflected in alpha1-frequency band activity. These data support changes in brain network activity, but are contradictory, and further investigation is needed. There are significant changes in alpha activity, which may be related to changes in thalamocortical gating activities.
Then event-related potential (ERP) EEG studies, which are few, report some discrete neural circuitry psychedelic effects, and the little evidence which is available suggests that exposure to psychedelic drugs impacts the relevant neural circuitry needed for specific cognitive tasks.
In a randomized, double-blind study, psilocybin, the preferential 5HT2A antagonist ketanserin, or psilocybin with ketanserin was administered acutely, during the completion of a facial recognition task psilocybin enhanced positive mood and attenuated recognition of negative facial expression, which was reflected in P300 wave form amplitude, positive > negative. A second study by the same group which investigated the spatiotemporal dynamics of a modal object completion task found psilocybin to attenuate the N170 amplitude, particularly apparent during the processing of incomplete objects, while slightly enhancing P100 component. The authors found the attenuated N170 over right extrastriate and posterior parietal cortices correlated with intensity of visual hallucinations. The authors suggest this reduction in N170 reflects 5HT1A and 5HT2A receptor-mediated visual hallucinations.
Then ERP cannabis studies report on occasional versus heavy cannabis users which completed a divergent attention task with acute administration of THC, occasional users showed reduced P100 amplitude; while both occasional and heavy users showed decreased P300 amplitudes, no effect on ERP waveforms were reported for their second task, the stop signal task, a task which measures activation of behavioral inhibitory circuitry. A second study, which addressed acute dose-related effects of THC on a three-stimulus oddball paradigm, found that with increasing dose of THC, P300a increased and P300b decreased, latency of P300 and wave-form of the N100 were not affected. The sparse ERP wave form findings suggest neural circuit function, versus state, is dependent on psychedelic exposure and dosing, beyond this further research needs to be conducted to gain better insight to the effects of psychedelics on neural processing during cognition.
THE MYSTERY
Wisdom requires not only the investigation of many things but contemplation of the mystery. -Jeremy Narby
Science is at the early stages of understanding psychedelic molecular mechanisms and even further from understanding the psychedelic state as any scientist who has experienced it is aware. Let us try to imagine how psychedelics are able to make conscious previously subconscious information, which forms part of traditional practitioners' imperative in administering psychedelics for therapeutic purposes over the millennia. This may provide insight to the relevant brain changes needed to promote attenuation of anxiety and promote anti-addictive brain states.
At a 2-month follow-up, approximately 70 of the participants rated the psilocybin experience as among the most personally meaningful of their lives. A subsequent study documented that administration of psilocybin led to increases in the open-ness domain of personality that was stable for at least a year. This was notable because few, if any, previous studies had demonstrated that any discrete experimental manipulation was capable of yielding long-lasting changes in personality. -Murnane
The 5-MeO-DMT state began with images of floating through the universe and being surrounded by the stars. He also described seeing a "universal cosmic matrix" that had a central column of electric light and spiritual beings merging into the light.
"It's just love. Everything. All of it. That is all that exists. Love is it. Upon debriefing from his session several hours afterwards, he believed this experience was the single-most peak transformational experience of his life. He reported he lost all sense of his body and surroundings, and was transformed on a cellular level into infinite energy and pure love, and described, all of [his] stress and difficulties throughout [his] life felt like they occurred for a meaningful purpose, and the traumas of [his] past were washed over by an infinitely loving energy." -Barsuglia et. al.
In individuals with substance use disorders, ibogaine stimulates heightened memory retrieval specifically related to drug abuse, the perception of one's own future with or without drug use, and visions which reveal powerful insights into the nature of the addiction such as personal traumas. -Barsuglia et. al.
Psychedelics are known to have exceptional anti-amnesic effects where memories are retrieved in great detail. In fact, all psychedelics have the ability to make conscious/reveal/retrieve that which was previously unconscious and the term, psychedelic, means mind-revealing.
The neurophysiological mechanisms that facilitate this anti-amnesic effect are largely unknown but progress is being made in identifying the molecular and cellular mechanisms and brain networks involved.
The anti-amnesic effect of psychedelics has also been supported in rodent studies where administration of ibogaine facilitated spatial memory retrieval, and low dose THC has been shown to improve memory in aged rodents, and to enhance synaptic marker proteins and increase hippocampal spine density. The anti-amnesic effect of ibogaine and ayahuasca is, partly, due to the SIGMAR1 affinity of both compounds. SIGMAR1 activation has been shown to reverse experimental-induced amnesia in rodents, via enhancement of the cholinergic and glutamatergic systems. Peak densities of SIGMAR1 are found in brain areas relevant to traumatic memory formation, retrieval and updating, such as the amygdala and the hippocampal formation.
Psychedelics also enhance synaptic plasticity and increase neurogenesis, processes known to be involved in memory reconsolidation and fear extinction. The fear response triggered by the memory can be reprogramed and/or extinguished through synaptic plasticity and changes in gene expression mediated by epigenetic modification via 5HT2A and SIGMAR1 activation.
Subsequently, the memory is reconsolidated and stored with updated significance via cortical mechanisms. As suggested, there is a change in cortical control from frontal brain areas to parietal brain areas and, in part, this is related to hippocampal network activity. For example, varying doses of PCP have been shown to change the functional connectivity (phMRI) between several brain areas, including the fronto-cortical and hippocampal brain regions. Atasoy et al. report that changes in brain activity occur in a frequency-specific manner with LSD and psilocybin, and that these changes in power-law components lead to an expansion of the repertoire of active brain states and the emergence of more complex brain dynamics which heightens information processing capabilities.
Perhaps a similar mechanism may be involved in ancestral communication.
Higher doses of ibogaine generate its psychoactive effects, including hallucinations and the facilitation of communion with the spirits of the ancestors in rites of passage. Ibogaine ingestion in a religious context allows a bonding across time and space between consumers and their ancestors and fellow community members through a shared common experience of a distinctive system of belief and consciousness. -Corkery
Ayahuasca and ibogaine are the psychedelics most commonly associated with ancestor communication. Ayahuasceros and Bwiti tribesman summon the ancestors during ceremony. The Basotho people of southern Africa use Boophone disticha, a psychedelic bulb for ancestor communication as well. This may be culturally-specific interpretation but an alternate hypothesis proposed here is that the same mechanisms that are involved in psychedelics' anti-amnestic properties, i.e., their ability to retrieve and reconsolidate memories via changes in gene expression and brain activity, are the same mechanisms involved in making conscious information in our DNA and inherited epigenetic information. Genetic memory is a field of psychology and epigenetic modification from ancestral experience and/or trauma is now known to be heritable and able to affect offspring for many generations afterward. More research is required but if changes to our mind, memories and personality are due to changes in gene expression and epigenetic mechanisms and if psychedelics are able to make conscious stored neural information that is coded by genes and epigenetics, then it may be possible for information stored in our DNA and in our epigenome to be made conscious and in the case of the epigenome, reconsolidated in the same way as memories are. This is not a new concept in the psychedelic community. Psychedelics are said to heal ancestral trauma. After spending many years researching ayahuasca in South America, Narby concluded that ayahuasca used by shamans in the Western Amazon affords them access to knowledge coded in DNA. As more and more research uncovers the molecular mechanisms of psychedelic neuro-science, this does not seem like an impossible idea. Early psychedelic use in South America and Africa may be the origin of modern ancestor worship.
https://www.researchgate.net/publication/328974974_An_introduction_to_psychedelic_neuroscience
Last edited: